Agarwood has been widely used as therapeutic perfumes, traditional medicine, religious purposes and aromatic food ingredient (Liu Y. et al., 2013). Some of the earliest known uses of agarwood were recorded in ancient literatures, religious scriptures and medical texts. The word “aloes” which means agarwood was found occurring in the Sanskrit poet, Kâlidâsa that can be dated back to c. 4th–5th century CE (Lee and Mohamed, 2016). Meanwhile, the use of agarwood in the prescription of traditional Chinese medicine of the same period had also been recorded. The Chinese medicine uses it as a natural sedative, pain reliever, digestive aid and carminative (Ye et al., 2016; Liu Y. Y. et al., 2017).
Agarwood has high demand throughout the world as a raw material for incense, perfume and medicine purposes, with Middle East and East Asia as the two major regions of consumption (Antonopoulou et al., 2010). As the wealth of the consumer countries has gradually increased in the recent decades, the market’s demand for agarwood started to exceed its supply. Global agarwood prices can be ranging from US$ 20 – 6,000 per kilogram for the wood chips depending on its quality or US$ 10,000 per kilogram for the wood itself (Abdin, 2014). In addition, the value of agarwood essential oil can be as high as US$ 30,000 per kilogram. The annual global market for agarwood has been estimated to be in the range of US$ 6 – 8 billion (Akter et al., 2013), yet a large number of the trades have not been recorded.
The indiscriminate harvesting of agarwood from natural habitats has seriously hampered natural regeneration of Aquilaria trees, thus threatening the survival of the species in the wild. In order to meet the high market demand yet to protect the species from extinction, mass plantations of Aquilaria trees have been established across the Asian countries to allow sustainable agarwood production (Azren et al., 2018). Since agarwood formation in natural environment is a very long process which can take up to 10 years, the development of effective induction technology has received a great attention as it is extremely crucial to ensure the stability of agarwood yield from the domesticated Aquilaria trees.
Naturally, agarwood formation is often linked to the physical wounding or damage of Aquilaria trees caused by thunder strike, animal grazing, pest and disease infestations (Rasool and Mohamed, 2016; Wu et al., 2017). These events expose the inner part of the trees toward pathogenic microbes, which elicit the defense mechanism of Aquilaria to initiate the resin production. This natural formation process of agarwood has greatly inspired the development of diverse artificial induction methods (Table 1). For example, many traditional induction approaches like nail in setting, holing, burning, trunk breaking and bark removal have adopted the concept of physically wound the trees (Mohamed et al., 2010; Azren et al., 2018). Although it is cost effective and requires only personnel with little or no scientific knowledge on agarwood, but these induction methods usually result in inferior quality and uncertain yield of agarwood.
Instead of relying on external stimuli to trigger plant responses, either by mechanical wounding or biological inoculum, some induction approaches have been developed to introduce signaling molecules directly and specifically into Aquilaria trees to initiate agarwood resin biosynthesis pathways (Liu Y. et al., 2013; Wu et al., 2017). Chemical inducers normally comprise of phytohormones, salts, minerals and biological-derived substances (Zhang et al., 2012; Liu Y. et al., 2013; Van Thanh et al., 2015). Besides, suitable delivery method is often developed together with the chemical formulations to ease the large-scale induction process, such as vessel equipped with transfusion needle (Yang et al., 2014c). To date, several induction approaches have been developed based on the chemical induction concept such as cultivated agarwood kit (CA-kit), the whole-tree agarwood inducing technique (Agar-Wit) and biologically agarwood-inducing technique (Agar-bit). CA-kit is a combined method based on physical wounding and chemical induction, where the inducing agent is applied into the Aquilaria tree via an aeration device inserted into the wound (Blanchette and Heuveling, 2009). This method results in satisfying yield and quality, but the procedures are in some way conventional. On the other hands, Agar-Wit is a transpiration-assisted chemical treatment to form an overall wound in the tree, where the preloaded inducer in a transfusion set is distributed via plant transpiration (Liu Y. et al., 2013). Through this method, a larger agarwood coverage area can be achieved, but unfortunately produces more decayed tissues. Similarly, Agar-bit method adopts the idea of distributing the inducing reagent by plant transpiration, except that the reagents are injected directly into the stems of the tree (Wu et al., 2017).
Through chemical induction approach, the time-consuming holing process can be minimized as less induction sites are needed to deliver the inducers throughout the plants via transpiration process. Properly formulated inducer was shown to be able to produce artificial agarwood with quality closely resembled to those obtained from natural source (Liu Y. et al., 2013). In spite of the fast results and high yields, the application of chemical inducers still poses skepticism of toxicity on both human and environment. More assessments on chemical inducers are required to test its effectiveness on fields and also to popularize its use. Chemical inducers are undoubtedly more suitable for mass production with easier quality control than biological inoculum, which is highly potential to substitute conventional induction methods and the use of biological inoculum in agarwood industry.
The main attraction of the agarwood industry is its extremely high market value. Yet, the price of agarwood is largely determined by its quality which is graded solely based on human experience from the age-old practices of each country. The unavailability of standard quality grading system can be due to the intricate appearance of the traded agarwood and personal interest. The currently adopted agarwood quality assessment in the market has been extensively reviewed by Liu Y. Y. et al. (2017). Recently, the metabolite analysis of agarwood has gained increasing attention as some studies showed that there is correlation of agarwood quality to its resin yield and metabolite constituents (Pasaribu et al., 2015; Liu Y. Y. et al., 2017). Many studies have been conducted to clarify the metabolite composition of agarwood obtained either from wild or artificially induced methods (Chen et al., 2012; Gao X. et al., 2014; Hashim et al., 2014). It was concluded that the composition of agarwood resin is mainly composed of the mixtures of sesquiterpenes and 2-(2-phenylethyl) chromones (PECs) (Naef, 2011; Chen et al., 2012; Subasinghe and Hettiarachchi, 2015; Figure 1). Meanwhile, the constituents of agarwood essential oil were shown primarily to be sesquiterpenoids (Fazila and Halim, 2012; Hashim et al., 2014; Jayachandran et al., 2014). Together, all of these major compounds and some low abundant volatile aromatic metabolites form the unique and fragrant-smelling property of agarwood.

The PEC derivatives, as other major fragrance constituents of agarwood are the important contributors to the sweet, fruity and long lasting scent of agarwood when it is burnt. These compounds can only be detected by supercritical carbon dioxide and solvent extraction methods but never present in the extract of hydrodistillation (Yoswathana, 2013; Jong et al., 2014). In comparison to the sesquiterpene constituents in agarwood, the types of PECs being determined by GC-MS are relatively limited. Structural studies revealed that all previously reported PECs in agarwood own the same basic skeleton (molecular weight: 250) and similar substituents, i.e., either hydroxy or methoxy groups (Mei et al., 2013). The percentage of 2(2-phenylethyl) chromone and 2-(2-4-methoxy-phenylethyl) chromone in the high grade agarwood such as kanankoh can be as high as 66.47 %, which is overwhelmingly higher than the lower-quality agarwood jinkoh that has only 1.5% (Ishihara et al., 1993). Furthermore, the presence of certain PEC derivatives in agarwood was proposed to be useful in the evaluation of the grading of agarwood products (Shimada et al., 1982). There are 17 types of chromone derivatives which are agarwood specific and potential marker for the purpose of authentication (Naef, 2011). The substituted chromones, such as agarotetrol and isoagarotetrol (Figure 3), were shown to have positive correlation with the quality of agarwood obtained in the market with some exceptions (Shimada et al., 1986).
The types and derivatives of major compounds in agarwood are extremely wide and diverse, indicating the miscellaneous fragrance properties of agarwood from different species and regional sources. The better insight of agarwood metabolites will definitely facilitate the identification of universally accepted biomarkers for agarwood grading. Since the publication of the comprehensive review of Naef (2011) regarding the major constituents of agarwood, new compounds continue to be discovered in the later studies (Wu et al., 2012a; Yang et al., 2014b; Wang et al., 2015). The number of discovered compounds in agarwood will certainly be further increased in the future.
The Biosynthesis Pathways of Agarwood Constituents
Agarwood formation can be related to the self-defense mechanism of Aquilaria trees in response to biotic and abiotic stresses (Gao et al., 2012b; Singh and Sharma, 2015). Stresses trigger the defense responses of Aquilaria species which in turn initiate the secondary metabolite biosynthesis and the accumulation of agarwood resin. Previously, we have mentioned that sesquiterpenes and PEC derivatives are the main constituents in agarwood. Hence, it is crucially important to understand the metabolic pathway for the regulation and biosynthesis of sesquiterpenes and chromone derivatives in Aquilaria species to effectively induce the agarwood formation.
In plants, the isoprenoid precursors for the biosynthesis of sesquiterpenes, triterpenes and sterols has generally been assumed to be provided from the mevalonic acid (MVA) pathway in cytosol. In plastids, the 1-deoxy-D-xylulose-5-phosphate (DXP) or known as methylerythritol phosphate (MEP) pathway provides precursors for the production of monoterpenes, diterpenes, and carotenoids (Rohmer, 1999; Dong et al., 2015; Singh and Sharma, 2015). These two pathways biosynthesise C5 homoallylic isoprenoid precursor, that is isopentenyl pyrophosphate (IPP) and its electrophilic allylic isomer dimethylallyl pyrophosphate (DMAPP). An exchange of IPP and DMAPP was observed to happen in between plastids and cytosol even with the spatial partitioning of the two pathways (Dong et al., 2015). The production of IPP and DMAPP precursors from pyruvate and acetyl-CoA involves a series of enzymes according to the respective pathway (Figure 4). The genes encode for these enzymes have been identified from Aquilaria species through transcriptome sequencing analysis (Xu et al., 2013; Ye et al., 2016). These C5 isoprene units will later be channeled into the generation of C15 farnesyl pyrophosphate (FPP) by sequential condensation reactions in the presence of FPP synthase (FPS) (Rohmer, 1999; Yang et al., 2013; Ye et al., 2016). The FPS is one of the key-limiting enzymes responsible for the sesquiterpene biosynthesis (Gaffe et al., 2000; Yang et al., 2013; Liu X. M.et al., 2017). The genes encode for FPS have been cloned from Aquilaria microcarpa (Am-FaPS-1) (Kenmotsu et al., 2011) and Aquilaria sinensis (AsFPS1) (Yang et al., 2013). The transcript level of AsFPS1 was reported to be higher in stem and roots than the leaves, suggesting that sesquiterpene synthesis in Aquilaria species tends to be tissue-specific. Besides, the expression of Am-FaPS-1 was shown to be up-regulated upon exposure to methyl jasmonate (MeJA), yeast extract and Ca2+-ionophore A23187, indicating that the two former chemicals are effective to initiate the sesquiterpene biosynthesis pathway whereas Ca2+ can act as signaling molecule during the activation process (Kenmotsu et al., 2011). This provides clues for the artificial induction of agarwood formation via exogenous chemically induced approaches by triggering the sesquiterpene biosynthetic pathway in Aquilaria trees.

FIGURE 4. Schematic relationships between the wound-induced signal transduction mechanisms for the sesquiterpene biosynthesis and regulation in Aquilaria species for the agarwood production. External stimuli trigger the Ca2+ signaling pathway and induce the defense responses of Aquilaria species via hydrogen peroxide (H2O2) pathway, ethylene (ET) signals, Jasmonic acid (JA) signals, and salicylic acid (SA) signals. MeJA treatment triggers H2O2 production that can induce programmed cell death (PCD) and increase the sesquiterpene synthesis. These signaling molecules activate the transcription factors such as MYB, MYC, and WRKY, which will bind to the cis-element on the promoter of terpenes biosynthesis genes in the mevalonic acid (MVA) and methylerythritol phosphate (MEP) pathways and also the downstream terpene synthase genes (TPs). Direct and indirect interactions are shown as solid and dotted lines, respectively. AACT, acetyl-CoA C-acetyl transferase; HMGS, hydroxymethylglutaryl (HMG)-CoA synthase; HMGR, HMG-CoA reductase; MK, mevalonate kinase; MPK, phosphomevalonate kinase; MDD, mevalonate diphosphate decarboxylase; DXP, 1-deoxy-D-xylulose 5-phosphate; DXS, DXP synthase; DXR, DXP reductoisomerase; CMK, 4-(cytidine 50-diphospho)-2-C-methyl-D-erythritol kinase; MCS, 2-C-methyl-D-D-erythritol-2,4-cyclo diphosphate synthase; HDS, (E)-4-hydroxy-3-methylbut-2-enyl diphosphate synthase; IDS, isopentenyl diphosphate synthase; GPS, geranyl disphosphate synthase; GGPS, geranylgeranyl diphosphate synthase.
In the final stage of sesquiterpenes production, the enzymes accountable for the diversification of sesquiterpene mainly come from the classes of sesquiterpene synthases (SesTPs) and cytochrome P450 dependent mono-oxygenases (P450s). The SesTP enzymes are responsible to catalyze the formation of multicyclic scaffold complexes from FPP, followed by oxidative functionalization of the resulting scaffolds by cytochrome P450 enzymes. The added hydroxyl groups by P450s can serve as molecular handles for further modifications, such as alkylations, esterifications and the addition of sugar residues (Pateraki et al., 2015). In addition, the P450 enzymes which carry out stereospecific hydroxylation on the hydrocarbon backbones, that is important for the novel chiralities and further modifications of the sesquiterpene molecules, have never been reported from Aquilaria species thus far. Similarly, the NADPH-dependent cytochrome P450 oxidoreductases (POR) in Aquilaria that act as redox partners of P450s catalysis activity are basically unexplored. Several studies have been reported to isolate genes encode for SesTPs from Aquilaria, which can be considered as the early attempts to study SesTPs involved in the agarwood formation (Kumeta and Ito, 2010; Xu et al., 2013). In the study of Kumeta and Ito (2010), five genes encode for sesquiterpene synthases which shared highly similar amino acid sequences have been isolated from Aquilaria crassna. Three out of these genes have been successfully expressed in Escherichia coli and enzymatically converted FPP into δ-guaiene as their major product. Besides, three other sesquiterpene synthase genes (ASS1, ASS2, and ASS3) identified from A. sinensis via transcriptome sequencing have been revealed to encode enzymes that produce δ-guaiene as well (Xu et al., 2013). The isolation of SesTP genes was also described in A. malaccensis where the temporal and spatial expression of the two SesTPs reported in the study, i.e., the guaiene (AmGuaiS1) and sesquiterpene synthase (AmSesTPS1), was elucidated (Azzarina et al., 2016). The AmSesTPS1 was found to be highly expressed after 6 h of wounding while AmGuaiS1 was induced after 2 h of wounding at a magnitude of 18- and 5.5-fold higher than unwounded control, respectively. Recently, a novel sesquiterpene synthase gene (As-sesTPS) was isolated from A. sinensis where the recombinant As-sesTPS catalyzed FPP into nerolidol (Ye et al., 2018). Expression analysis showed that the transcript level of As-sesTPS was much higher in agarwood than the healthy wood, implying that the gene can be participated in the agarwood formation. Despite the fact that many sesquiterpene compounds have been discovered from the agarwood, the corresponding SesTPs responsible for their production have yet to be reported from Aquilaria. For that reason, the sesquiterpene biosynthesis pathways involve in the functionalization of terpenes in Aquilaria is urged for further clarification.
On the other hand, chromones are a large group of secondary metabolites with wide-ranging potential therapeutic indications toward immunomodulation, inflammation, cancer, diabetes, neurological conditions, bacterial and viral infections (Khadem and Marles, 2011; Yang et al., 2012; Tawfik et al., 2014). Chromone is derived from a polycyclic organic compound namely benzopyran ring, with a keto group substitution on its oxime ring. It is generally believed that derivations of chromones take place as a consequence of the convergence of multiple secondary metabolite biosynthetic pathways involving pentaketide pathway, shikimic acid pathway and the addition of nitrogenous moiety from amino acids or other sources (Khadem and Marles, 2011). Owing to the extensive pharmacological properties associated with its bicyclic ring structure, chromones have been used as the privileged scaffold in the development of new drugs (Reis et al., 2017). The PECs are small class of chromones, which hold a phenylethyl substituent at the C2 of benzopyran ring of the chromone that happened to be structurally unique in the family (Ibrahim and Mohamed, 2015). Until now, the PECs have only been found to be present in a few species of plants for example Bothriochloa ischaemum (Wang et al., 2001), Imperata cylindrical (Liu X. et al., 2013), Cucumis melo L. (Ibrahim, 2014), Gyrinops salicifolia (Shao et al., 2016), and Aquilaria species (Wu et al., 2012b; Yang et al., 2014a). Recently, a hypothetical scheme for the biosynthetic pathway of PECs was proposed by Liao et al. (2018) based on in-dept analysis of agarwood chemical constituents using GC-EL-MS and UPLC-ESI-MS/MS methods. In their study, the PECs was found to be the major agarwood resin constituents, which is comprised mostly of flindersia-type 2-(2-phenylethyl) chromones (FTPECs). The formation of FTPECs is further elucidated to be possibly catalyzed by type III polyketide synthase (PKs) through condensation of dihydro-cinnamoyl-CoA analogs and malonyl-CoA with 2-hydroxy-benzoyl-CoA to produce PEC scaffold that will subsequently be catalyzed by hydroxylases or O-methyltransferases (OMTs) to form structurally diverse FTPECs (Liao et al., 2018). Recent study showed that salinity stress could induce the biosynthesis of PECs in A. sinensis calli (Wang et al., 2016). Transcriptomic analysis of these salt-induced A. sinensis calli have identified several upregulated candidate genes potentially involved in the biosynthesis of PECs, including three OMT-encoding genes (flavonol-OMT 1, flavonol-3-OMT and caffeoyl-CoA-OMT) and a type III polyketide synthase gene encodes for chalcone synthase 1 (AsCHS1).
In spite of the recent progress made on the understandings of PECs biosynthesis, a tremendous effort is necessary to experimentally determine the missing steps in this complex PEC biosynthetic pathway. Even with the inadequate knowledge on the detailed PEC biosynthetic pathway, artificial synthesis of chromones and some of its derivatives is nevertheless feasible due to the advancement of chemical processes (Goel and Makrandi, 2006; Tawfik et al., 2014). Agarwood is a rich source of PEC derivatives which deserves further investigation to uncover the structure of new chromone compounds and improve the understanding toward its biosynthetic mechanism at the molecular level.
Signaling and Regulation Mechanism of Agarwood Formation
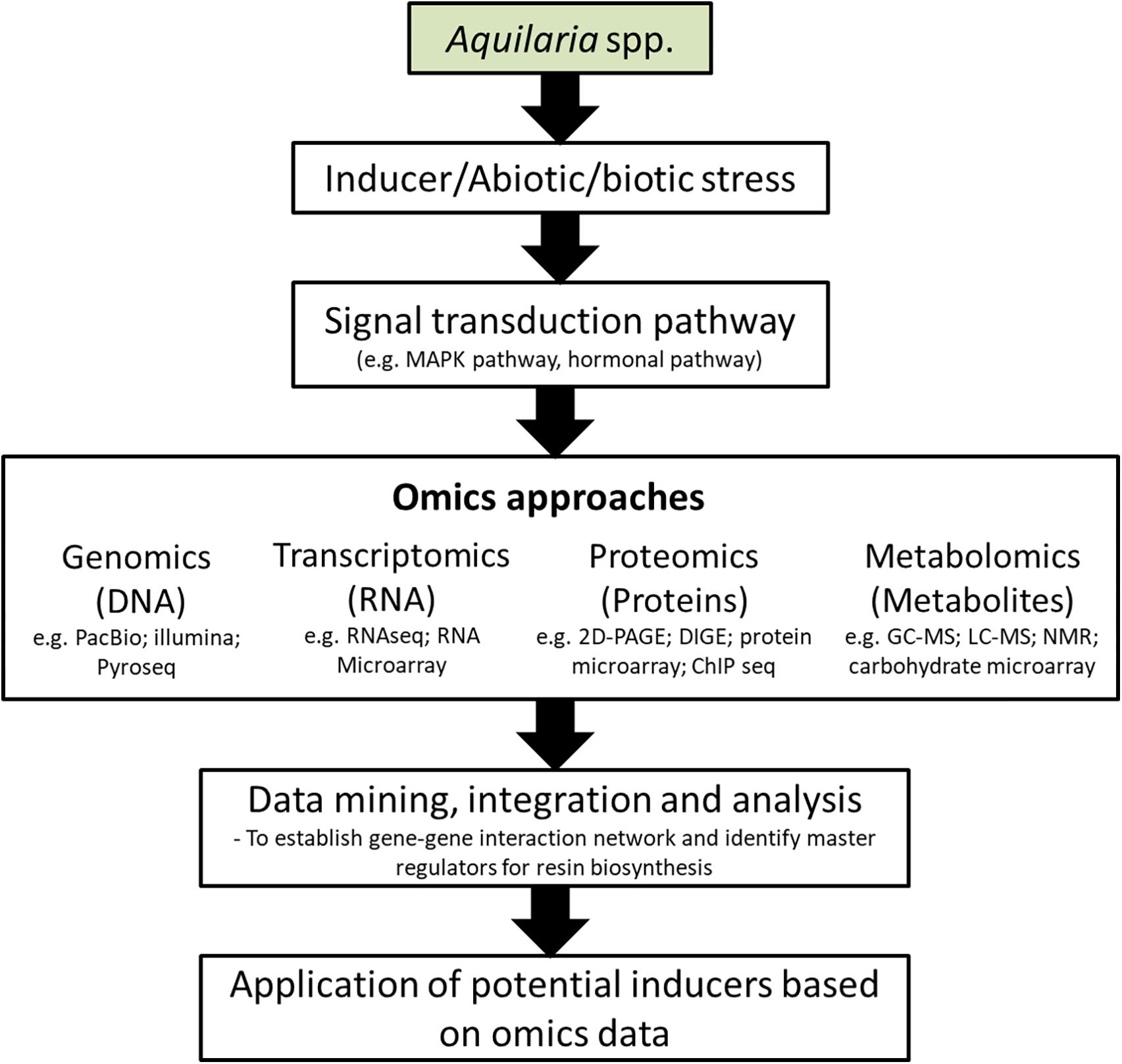
In nature, the probability of getting agarwood-containing Aquilaria trees are extremely low (1–2%), where can only be found on pathogenically infected or wounded trees (Cui et al., 2013; Chhipa and Kaushik, 2017). Therefore, it is sensible to assume that there is a wound-inducible signal transduction process causing the expression of sesquiterpene synthases prior to agarwood formation. In order to clarify the relationship of wound signal transduction and regulation of agarwood formation, high-throughput studies on agarwood formation have recently gained attention in researches (Table 2). A schematic diagram of the proposed signal transduction mechanism of sesquiterpene biosynthesis and regulation in Aquilaria species is provided in Figure 4.
The mitogen-activated protein kinase (MAPK) signaling pathway has been proposed as wound-induced signaling mechanism for the agarwood formation in A. sinensis, which phosphorylates downstream transcription factors (TFs) like MYB or WRKY that eventually lead to the expression of sesquiterpene synthase genes (ASSs) (Xu et al., 2013). The MAPK signaling cascade consists of three sequentially activated components [MAPK kinase kinases (MAPKKKs), MAPK kinases (MAPKKs), and MAPKs], which is highly conserved signaling mechanism in eukaryotes in mediating extracellular signals to downstream responsive genes (Sinha et al., 2011; Xu C. et al., 2017). Xu et al. (2013) has reported that a total of 41 unigenes from the transcriptome analysis of wounded A. sinensis are annotated as being related to MAPK signaling pathway and 25 to calcium signaling pathways which may play roles in wound-induced agarwood formation. In plants, calcium ions (Ca2+) are important intracellular secondary messenger molecules to regulate many signal transduction pathways reacting to the external stimuli (Tuteja and Mahajan, 2007). Earlier studies in other plants have shown that TFs are crucial regulators in stress-responsive signaling pathways to transmit signals to different cellular centers to activate plant adaptation/defense mechanisms against adverse environments, including TFs like bZIP, ERF, EIN3, MYB, MYC, and WRKY (Ambawat et al., 2013; Phukan et al., 2016; Schmiesing et al., 2016). Overexpression of AaWRKY1 in Artemisia annua was found to have positively regulated the expression of amorpha-4,11-diene synthase gene (ADS) and significantly increased the production of artemisinin (Ma et al., 2009). The expression of cotton TF GaWRKY was also shown to upregulate the sesquiterpene synthase genes for the biosynthesis of (+)-δ-cadinene and gossypol sesquiterpene (Xu et al., 2004).
Besides mechanical wounding, the MeJA is an effective elicitor to increase the sesquiterpenes content in Aquilaria (Xu et al., 2013, 2016; Xu Y. H. et al., 2017). Previous studies have shown that heat shock can increase the expression of genes involved in the Jasmonic acid (JA) biosynthesis, including allene oxide cyclase (AOC), allene oxide synthase (AOS), lipoxygenase (LOX) and 12-oxophytodienoate reductase 3 (OPR3) genes, which subsequently lead to the production of JA and the accumulation of sesquiterpene in the A. sinensis suspension cell culture (Xu et al., 2016). The A. sinensis coronatine-insensitive protein 1 (AsCOI1), acts as a receptor in MeJA signaling pathway, has been cloned and characterized (Liao et al., 2015). Expression study of AsCOI1 has demonstrated that the gene was expressed in a tissue-specific pattern which is highest in stem, followed by root and leaves. The findings imply that resin production due to external stimuli may be more responsive in stem of Aquilaria. The AsCOI1 gene is responsive toward early treatment of MeJA, mechanical wounding and heat stress. The application of MeJA in A. sinensis has increased the expression level of 17 wound signaling-related genes, including TFs WRKY4 and MYB4, protein kinases CAPKs, MAPKs and MAPKK, NADPH oxidase noxB and some regulators related to signal molecules MeJA, ethylene and hydrogen peroxide (Xu et al., 2013). Interestingly, the hydrogen peroxide (H2O2) producing NADPH oxidase noxB was found to be significantly up-regulated by the MeJA treatment (Xu et al., 2013; Gong et al., 2017), which is consistent with the findings that MeJA triggers H2O2 production in plants (Orozco-Cardenas et al., 2001; Hung et al., 2006). A study on suspension cultures of A. sinensis revealed that H2O2 can induce the programmed cell death (PCD) and sesquiterpene synthesis by the elevated expression of ASS genes due to the endogenous accumulation of salicylic acid (SA) (Liu et al., 2015). Meanwhile, the expression of jasmonate-responsive key sesquiterpene synthase ASS1 in A. sinensis was described to be regulated by a TF AsMYC2 (Xu Y. H. et al., 2017). As an immediate-early responsive gene toward MeJA treatment, AsMYC2 binds to the ASS1 promoter containing a G-box motif to initiate the expression of ASS1. Similarly, the homolog of AsMYC2 in Arabidopsis (MYC2) was shown to be MeJA-responsive and up-regulated the expression of two sesquiterpene synthases (TPS11 and TPS21) upon treatment (Hong et al., 2012). In addition, MeJA treatment has successfully induced the synthesis of 3 sesquiterpenes, that are α-guaiene, α-humulene and δ-guaiene in A. crassna cell culture (Ito et al., 2005; Kumeta and Ito, 2010).
In contrary to the sesquiterpene biosynthesis pathway, the biosynthesis and regulation of PECs remains almost unknown. In the study of Wang et al. (2016), forty-one PECs have been produced in A. sinensis calli by salt treatment. With the help of high-throughput transcriptome analysis, a total of 18,069 differentially expressed transcripts between the control and the NaCl-treated A. sinensis calli induced by 24 or 120 h salinity stress were obtained. Many differentially expressed genes are reported to be involved in the hormone signal transduction; including genes encode for MAPK cascades, receptor-like kinases, Ca2+ signal transduction and transcription factors (Wang et al., 2016).
Nonetheless, the pace of research did not just stop at searching for effective inducers and their underlying signal transduction process, but also extended to their post-transcriptional regulation mechanism. By deep sequencing of sRNAs from healthy control and wounded samples of A. sinensis, Gao et al. (2012a) have identified ten stress-responsive miRNAs from 74 putative conserved miRNAs and their hairpin forming precursors were also confirmed. Expression pattern revealed that six of these stress-responsive miRNAs were up-regulated, including miR159, miR168, miR171, miR396, miR397, and miR408, whereas miR160 and miR398 were down-regulated and continued their reduced level at 2 day point (Gao et al., 2012a). The different responses of miRNAs reacted toward treatment and the effect lasted for various time lengths reflecting the diversity of their positions in the post-transcriptional regulation of wound response in A. sinensis. Among the identified miRNAs, the down-regulated miR398 is of interest where it was demonstrated to negatively regulate the pathogen-associated molecular pattern (PAMP)-triggered callose deposition and plant innate immunity against bacteria (Li et al., 2010). The oppositely reacted miR160 and miR398 in A. sinensis suggested that they might be important regulators and play a more distinct role on agarwood formation.
Further study on miRNAs profiling of wounded A. sinensis showed that some of the most conserved miRNAs such as miR159 and miR396 families elevated and subsided quickly in early period of treatment implying their function at the upstream of wound responses (Gao Z. H. et al., 2014). The down-regulated miR396b2 in wounded tissues of A. sinensis was suggested to involve in the biosynthesis and accumulation of agarwood constituents. The target of miR396b2, which has glutamyl-tRNA reductase activity, was believed to bind NADP and produce NADPH. The NADPH is the cofactor for the two key enzymes in terpene biosynthesis, i.e., 1-deoxy-D-xylulose 5 phosphate (DXR) and 3-hydroxy-3-methylglutaryl-CoA (HMGR) (Nagegowda, 2010). Furthermore, plant P450s that oxidatively functionalize the terpene scaffolds also require reducing agents for its catalytic activity, which is commonly provided by NADH or NADPH. Despite in the situation of lacking sequenced genome, high-throughput transcriptome analysis provides a feasible approach to examine the overall changes of gene expression on Aquilaria species responding to a variety of stresses. Further investigation of the functions of the identified regulator sequences would help to reveal the regulation mechanism of agarwood formation.
Future Prospects of Agarwood Induction Technology
Previous studies have shown that agarwood formation can be influenced by many factors. Together with the exceedingly complex agarwood resin composition, it is believed that agarwood formation is an intricate process which involved a variety of physiological changes occurs on Aquilaria trees to cope with the external stimuli either in the form of biotic or abiotic. This whole agarwood formation process is in any case inseparable from the gene-expression response of the trees toward the triggering factors. Consequently, future improvement of agarwood induction technology should emphasize on two aspects that are to further improve the induction efficiency and to screen more responsive lines of Aquilaria for resin production under breeding program.
In order to improve the induction efficiency, an induction technique plays a decisive role. The concept of currently available induction approaches can be summarized as either to provide external stimuli to activate the production of plant signaling molecules that eventually lead to the resin biosynthesis, or to bypass the external stimuli via direct introduction of signaling molecules to the plants. In any case, the overall concern is to increase agarwood yield and quality as well to reduce human intervention (e.g., holing process) during the induction process. Since the aforementioned physical wounding and biological induction method have their inevitable drawbacks of inconsistent agarwood quality and requiring intensive workforce, the chemical induction method can be regarded as a promising approach for further optimization (Table 1). Comprehensive understanding of agarwood formation at the molecular level via high-throughput using omics approach such as trancriptomic and metabolomic appears to be advantageous for more targeted and directional improvement of the induction formulation rather than the trial- and error-based experimentations.
By coupling with omics approaches such as single molecule real time sequencing technology (SMRT) which offers longer read lengths and highly contiguous de novo assemblies (Rhoads and Au, 2015), thus it tends to be particularly useful for unsolved problems in genome and transcriptome of non-model Aquilaria species where their genomes are not available. With longer reads, the highly repetitive non-coding regulatory sequence of genes controlling the agarwood resin production can be easily obtained. Sequence similarity/identity-based integration can be used to establish the cross-reference data sets between the query sequences and their homologous references from various public databases and genetic resources, where genomic sequence structures, domain features, promoter regions and gene ontology for motifs can be assigned (Mochida and Shinozaki, 2011). The development of sequencing technology has made the future research on the whole genome sequencing of Aquilaria species to be easier to fill in the lack of genome-wide information in the current situation. Data integration based on genome sequence is important to allow analysis of global changes of transcriptome through whole genome microarrays. The gene expression analysis of induced Aquilaria at its entirety can be examined in broad coordinated trends by this approach, which is indiscernible by individual assays. The expression profiles obtained in this way will help to identify potential agarwood-producing biomarker genes that are important indicators for downstream applications of agarwood induction.
The integration of systems biology and omics approaches, covering genomics, transcriptomics, proteomics, metabolomics and functional analysis; provide a potential solution to comprehend the multigenic nature of resin biosynthesis in Aquilaria. On the basis of large number of previously conducted agarwood induction experiments, transcriptomic and metabolites studies (Naef, 2011; Gao X. et al., 2014; Ye et al., 2016; Wu et al., 2017), integrated omics analysis can indeed serve as a platform to build a more comprehensive picture of agarwood resin biosynthetic mechanism that involve different omics layers. The development of high-throughput mass spectrometry (MS), microarray and sequencing technologies (DNA and RNA) have made it possible to integrate such data into a system biological framework via integrated-omics which would help to predict gene-gene interactions, identify driver genes and the molecular signatures of agarwood formation (Figure 5). Likewise, potential signaling molecules for agarwood formation can be predicted in a wake of this dramatically increased availability of data. Experimentally validated effective signaling molecules can be added into the existing recipe of chemical inducer to further enhance the induction performance. Moreover, predetermination of resin composition or the agarwood quality is conceivable with deep understanding of the key quality indicators and the specific pathways involved for agarwood production by integrated elucidation of different metabolite and transcript profiles under different induction methods. Future improvement of agarwood induction efficiency should accompany with the development of monitoring system for early detection of non-responsive trees which could avoid cutting down of Aquilaria trees that are unsuccessfully been induced. This can be achieved by monitoring the expression of a set of genes involved in agarwood resin biosynthesis.
Aside of the effective inducer, the responsiveness of Aquilaria trees toward stimulation is another determining factor for the production of agarwood. By knowing that the degree of plant response on stimuli is largely dependent on their genetic makeup, the utilization of highly responsive Aquilaria line as induction target is expected to further increase the agarwood yield rather than optimizing the inducer recipe alone. Conventionally, selective breeding based on phenotypic selection have been adopted to develop new plant lines with desirable traits. Current technology associates these beneficial traits of plants to genetic (DNA/RNA variations) or biochemical (signature metabolites) markers to allow marker-assisted selection (MAS). The approach of MAS offers a great promise for the selection of elite Aquilaria lines as these biomarkers can be applied to predict the phenotypic characteristics before these features develop into more noticeable. Such biomarkers can also be used for the development of fast and targeted diagnostic assays that will assist the selection program. As an alternative to obtain a high-yield line, a combined approach of genetic engineering (e.g., CRISPR-Cas9 genome editing technology) with tissue culture could pose a possibility to manipulate the key regulator genes of Aquilaria involved in the agarwood production which will help to fine tune or redirect the metabolic flux toward desired metabolic pathways. On the whole, an integrated and high-throughput strategy will provide sufficient information to continually improve the agarwood induction methods, which is superior compare to the traditional way of induction method establishment that rely on the visual observation and personal experience. A deeper insight into the essential compounds and the biosynthesis mechanism of agarwood resin would greatly ease to control the stability of agarwood yield, quality and its price in the future.
Trầm hương là phần nhựa của cây dó bầu ngoài gỗ, là sản phẩm có giá trị cao về y học và hương liệu. Để bảo vệ các loài Aquilaria có nguy cơ tuyệt chủng, việc trồng cây Aquilaria hàng loạt đã trở thành một cách bền vững ở các nước châu Á để thu được loại trầm hương có giá trị cao. Vì chỉ cây Aquilaria được kích hoạt về mặt sinh lý mới có thể tạo ra trầm hương, nên các phương pháp cảm ứng hiệu quả đã được tìm kiếm từ lâu trong ngành công nghiệp trầm hương.
Trầm hương (còn được gọi là gaharu ở Đông Nam Á, oud ở Trung Đông, chen xiang ở Trung Quốc, jinkoh ở Nhật Bản và agar ở Ấn Độ) là một loại gỗ có lõi nhựa thơm sẫm màu có giá trị cao của loài Aquilaria (Liu Y. Y. et al., 2017 ). Sự hình thành trầm hương thường liên quan đến vết thương và nhiễm nấm trên cây Aquilaria (Liu Y. et al., 2013; Mohamed et al., 2014). Nhựa được cây tiết ra như một phản ứng tự vệ và lắng đọng xung quanh vết thương trong nhiều năm sau vết thương, nơi mà sự tích tụ của các hợp chất dễ bay hơi cuối cùng tạo thành trầm hương (Subasinghe và Hettiarachchi, 2013).
Trầm hương đã được sử dụng rộng rãi làm nước hoa trị liệu, y học cổ truyền, mục đích tôn giáo và nguyên liệu thực phẩm thơm (Liu Y. et al., 2013). Một số công dụng sớm nhất được biết đến của trầm hương đã được ghi lại trong các tài liệu cổ xưa, kinh sách tôn giáo và văn bản y học. Từ “lô hội” có nghĩa là trầm hương đã được tìm thấy ở nhà thơ tiếng Phạn, Kâlidâsa có thể có từ c. thế kỷ thứ 4–5 CN (Lee và Mohamed, 2016). Trong khi đó, việc sử dụng trầm hương trong đơn thuốc y học cổ truyền Trung Quốc cùng thời cũng đã được ghi nhận. Y học Trung Quốc sử dụng nó như một loại thuốc an thần tự nhiên, giảm đau, hỗ trợ tiêu hóa và tống hơi (Ye et al., 2016; Liu Y. Y. et al., 2017).
Trầm hương có nhu cầu cao trên khắp thế giới như một nguyên liệu thô cho mục đích làm hương, nước hoa và y học, với Trung Đông và Đông Á là hai khu vực tiêu thụ chính (Antonopoulou et al., 2010). Khi sự giàu có của các quốc gia tiêu dùng tăng dần trong những thập kỷ gần đây, nhu cầu của thị trường về trầm hương bắt đầu vượt quá khả năng cung cấp. Giá trầm hương toàn cầu có thể dao động từ 20 – 6.000 USD/kg đối với dăm gỗ tùy thuộc vào chất lượng của nó hoặc 10.000 USD/kg đối với gỗ (Abdin, 2014). Ngoài ra, giá trị của tinh dầu trầm hương có thể lên tới 30.000 USD/kg. Thị trường trầm hương toàn cầu hàng năm được ước tính vào khoảng 6 – 8 tỷ đô la Mỹ (Akter et al., 2013), tuy nhiên một số lượng lớn các giao dịch vẫn chưa được ghi nhận.
Aquilaria thuộc họ thực vật hạt kín Thymelaeaceae, là loài đặc hữu của vương quốc Indomalaya. Cho đến nay, có tổng cộng 21 loài Aquilaria đã được ghi nhận và 13 loài trong số đó được công nhận là loài sản xuất trầm hương (Lee và Mohamed, 2016). Tuy nhiên, việc khai thác trầm hương một cách hủy diệt đã ảnh hưởng xấu đến quần thể hoang dã của tất cả các loài Aquilaria. Do đó, chi này hiện được liệt kê là loài có nguy cơ tuyệt chủng và được bảo vệ theo quy định của Công ước về buôn bán quốc tế các loài động vật và thực vật hoang dã có nguy cơ tuyệt chủng (CITES) do sự suy giảm nghiêm trọng của các loài trong tự nhiên (Công ước về buôn bán quốc tế các loài có nguy cơ tuyệt chủng [CITES], 2004; Lee và Mohamed, 2016). Nhu cầu trầm hương chất lượng cao cùng với sự cạn kiệt của các loài cây dó bầu hoang dã cho thấy giá trầm hương sẽ tiếp tục tăng cao. Thay vào đó, việc canh tác hàng loạt và trồng cây Aquilaria với quy mô lớn, đóng vai trò là nguồn bền vững để thu được trầm hương đã giải quyết rất nhiều tình trạng thiếu nguồn cung trầm hương trên thị trường toàn cầu.
Vì cây Aquilaria khỏe mạnh không tạo thành trầm hương, nên nó gần như không có giá trị gì, sự khan hiếm trầm hương tự nhiên đã thúc đẩy sự phát triển của các phương pháp tạo trầm hương nhân tạo. Những nỗ lực để tạo ra sự hình thành trầm hương một cách nhân tạo có thể được bắt nguồn từ đầu năm 300 sau Công nguyên trong lịch sử Trung Quốc, nơi người ta ghi lại rằng sự lắng đọng nhựa đi kèm với sự thay đổi màu sắc của các mô bên trong có thể xảy ra trong vòng một năm bằng cách làm tổn thương cây (López-Sampson và Trang, 2018). Bên cạnh phương pháp tạo vết thương cơ học, việc sử dụng các kỹ thuật gây bệnh bằng hóa chất, côn trùng và mầm bệnh ngày càng phổ biến trong ngành công nghiệp trầm hương hiện nay (Liu Y. và cộng sự, 2013; Mohamed và cộng sự, 2014; Kalita, 2015). Tất cả các kỹ thuật cảm ứng này trong mọi trường hợp đều bắt chước các quá trình hình thành trầm hương tự nhiên, có điểm mạnh và điểm yếu riêng. Trong bài viết này, chúng tôi cố gắng cung cấp một cái nhìn toàn diện hơn về các phương pháp cảm ứng hiện có và triển vọng phát triển của chúng bằng cách sử dụng sự tiến bộ của công nghệ sinh học. Để hiểu rõ hơn về quá trình hình thành trầm hương, cơ chế phân tử của các con đường sinh tổng hợp chất chuyển hóa thứ cấp làm cơ sở cho quá trình sản xuất nhựa cũng sẽ được trình bày chi tiết.
Phương pháp cảm ứng trầm hương
Việc khai thác trầm hương bừa bãi từ môi trường sống tự nhiên đã cản trở nghiêm trọng quá trình tái sinh tự nhiên của cây Aquilaria, do đó đe dọa sự tồn tại của loài này trong tự nhiên. Để đáp ứng nhu cầu cao của thị trường nhưng vẫn bảo vệ loài này khỏi nguy cơ tuyệt chủng, các đồn điền cây Aquilaria hàng loạt đã được thiết lập trên khắp các quốc gia châu Á để cho phép sản xuất trầm hương bền vững (Azren et al., 2018). Do quá trình hình thành trầm hương trong môi trường tự nhiên là một quá trình rất dài, có thể mất đến 10 năm, nên việc phát triển công nghệ cảm ứng hiệu quả đã nhận được sự quan tâm lớn vì nó cực kỳ quan trọng để đảm bảo sự ổn định của sản lượng trầm hương từ cây Aquilaria thuần hóa.
Đương nhiên, sự hình thành trầm hương thường liên quan đến vết thương hoặc thiệt hại vật lý của cây Aquilaria do sét đánh, động vật chăn thả, sâu bệnh (Rasool và Mohamed, 2016; Wu và cộng sự, 2017). Những sự kiện này khiến phần bên trong của cây tiếp xúc với các vi khuẩn gây bệnh, chúng gợi ra cơ chế bảo vệ của Aquilaria để bắt đầu sản xuất nhựa. Quá trình hình thành trầm hương tự nhiên này đã truyền cảm hứng rất lớn cho sự phát triển của các phương pháp cảm ứng nhân tạo đa dạng (Bảng 1). Ví dụ, nhiều phương pháp cảm ứng truyền thống như đóng đinh, đục lỗ, đốt, bẻ thân và loại bỏ vỏ cây đã áp dụng khái niệm gây vết thương vật lý cho cây (Mohamed và cộng sự, 2010; Azren và cộng sự, 2018). Mặc dù tiết kiệm chi phí và chỉ yêu cầu nhân sự có ít hoặc không có kiến thức khoa học về trầm hương, nhưng các phương pháp cảm ứng này thường dẫn đến chất lượng trầm hương kém hơn và sản lượng trầm hương không ổn định.
Với sự hiểu biết nhiều hơn về các tương tác Aquilaria-nấm trong việc thúc đẩy sự hình thành trầm hương, các phương pháp cảm ứng dần dần chuyển từ vết thương cơ học đơn thuần sang vết thương có chủ ý kết hợp với việc áp dụng chất cấy sinh học (Jong và cộng sự, 2014). Nhiều chủng nấm nuôi cấy thuần khiết được phân lập từ trầm hương tự nhiên đã được phát hiện là tác nhân sinh học hiệu quả để tạo ra trầm hương ở những cây Aquilaria khỏe mạnh (Cui et al., 2013; Siburian et al., 2015; Sangareswari Nagajothi et al., 2016). Những cây Aquilaria bị nhiễm nấm đã được báo cáo là lắng đọng nhựa trầm hương xung quanh các vị trí bị nhiễm bệnh như một rào cản để ngăn chặn sự xâm nhập của nấm (Cui et al., 2013; Rasool và Mohamed, 2016). Một lợi thế rõ ràng của việc sử dụng chất cấy nấm là nó thường được cho là an toàn để xử lý và thân thiện với môi trường. Tuy nhiên, việc cấy nấm thường sẽ dẫn đến chất lượng trầm hương cục bộ và không đồng đều do các nhóm nấm khác nhau được sử dụng. Như một giải pháp, quy trình đục lỗ tốn nhiều công sức và thời gian ủ dài là cần thiết để tối đa hóa diện tích bề mặt bị xâm lấn trên cây để tạo ra chất lượng trầm hương tốt hơn (Mohamed và cộng sự, 2014).
Thay vì dựa vào các kích thích bên ngoài để kích hoạt phản ứng của cây, bằng cách gây tổn thương cơ học hoặc cấy sinh học, một số phương pháp cảm ứng đã được phát triển để đưa các phân tử tín hiệu trực tiếp và cụ thể vào cây Aquilaria để bắt đầu quá trình sinh tổng hợp nhựa trầm hương (Liu Y. et al., 2013 ; Wu và cộng sự, 2017). Các chất cảm ứng hóa học thường bao gồm phytohormone, muối, khoáng chất và các chất có nguồn gốc sinh học (Zhang et al., 2012; Liu Y. et al., 2013; Van Thanh et al., 2015). Bên cạnh đó, phương pháp phân phối phù hợp thường được phát triển cùng với các công thức hóa học để giảm bớt quá trình cảm ứng quy mô lớn, chẳng hạn như bình được trang bị kim truyền (Yang và cộng sự, 2014c). Cho đến nay, một số phương pháp cảm ứng đã được phát triển dựa trên khái niệm cảm ứng hóa học như bộ dụng cụ tạo trầm hương nhân tạo (CA-kit), kỹ thuật tạo trầm hương nguyên cây (Agar-Wit) và kỹ thuật tạo trầm hương sinh học (Agar-bit). CA-kit là một phương pháp kết hợp dựa trên vết thương vật lý và cảm ứng hóa học, trong đó chất gây cảm ứng được đưa vào cây Aquilaria thông qua một thiết bị sục khí được đưa vào vết thương (Blanchette và Heuveling, 2009). Phương pháp này dẫn đến năng suất và chất lượng đáp ứng, nhưng các quy trình theo một cách nào đó là thông thường. Mặt khác, Agar-Wit là một phương pháp xử lý hóa học hỗ trợ thoát hơi nước để tạo thành một vết thương tổng thể trên cây, nơi chất cảm ứng nạp sẵn trong bộ truyền được phân phối thông qua quá trình thoát hơi nước của cây (Liu Y. et al., 2013). Thông qua phương pháp này, có thể đạt được diện tích bao phủ trầm hương lớn hơn, nhưng không may là tạo ra nhiều mô bị phân hủy hơn. Tương tự, phương pháp Agar-bit áp dụng ý tưởng phân phối thuốc thử cảm ứng bằng cách thoát hơi nước của thực vật, ngoại trừ việc thuốc thử được tiêm trực tiếp vào thân cây (Wu et al., 2017).
Thông qua phương pháp cảm ứng hóa học, quá trình đục lỗ tốn thời gian có thể được giảm thiểu vì cần ít vị trí cảm ứng hơn để đưa chất cảm ứng đi khắp cây thông qua quá trình thoát hơi nước. Chất cảm ứng có công thức phù hợp đã được chứng minh là có thể tạo ra trầm hương nhân tạo với chất lượng gần giống với trầm hương thu được từ nguồn tự nhiên (Liu Y. và cộng sự, 2013). Mặc dù cho kết quả nhanh và năng suất cao, việc áp dụng chất cảm ứng hóa học vẫn đặt ra những nghi ngờ về độc tính đối với cả con người và môi trường. Cần có nhiều đánh giá hơn về chất cảm ứng hóa học để kiểm tra hiệu quả của nó trên các lĩnh vực và cũng để phổ biến việc sử dụng nó. Chất cảm ứng hóa học chắc chắn phù hợp hơn cho sản xuất hàng loạt với việc kiểm soát chất lượng dễ dàng hơn so với chất cấy sinh học, có tiềm năng cao để thay thế các phương pháp cảm ứng thông thường và việc sử dụng chất cấy sinh học trong ngành công nghiệp trầm hương.
Thành phần chính của trầm hương
Điểm hấp dẫn chính của ngành công nghiệp trầm hương là giá trị thị trường cực cao của nó. Tuy nhiên, giá của trầm hương phần lớn được quyết định bởi chất lượng của nó, được phân loại chỉ dựa trên kinh nghiệm của con người từ các tập quán lâu đời của mỗi quốc gia. Việc không có hệ thống phân loại chất lượng tiêu chuẩn có thể là do hình thức phức tạp của trầm hương được giao dịch và lợi ích cá nhân. Đánh giá chất lượng trầm hương hiện đang được áp dụng trên thị trường đã được xem xét rộng rãi bởi Liu Y. Y. et al. (2017). Gần đây, phân tích chất chuyển hóa của trầm hương ngày càng được chú ý vì một số nghiên cứu cho thấy có mối tương quan giữa chất lượng trầm hương với sản lượng nhựa và thành phần chất chuyển hóa (Pasaribu et al., 2015; Liu Y. Y. et al., 2017). Nhiều nghiên cứu đã được thực hiện để làm rõ thành phần chất chuyển hóa của trầm hương thu được từ phương pháp tự nhiên hoặc nhân tạo (Chen và cộng sự, 2012; Gao X. và cộng sự, 2014; Hashim và cộng sự, 2014). Người ta kết luận rằng thành phần của nhựa trầm hương chủ yếu bao gồm hỗn hợp sesquiterpenes và 2-(2-phenylethyl) chromones (PECs) (Naef, 2011; Chen et al., 2012; Subasinghe và Hettiarachchi, 2015; Hình 1) . Trong khi đó, thành phần của tinh dầu trầm hương chủ yếu là sesquiterpenoids (Fazila và Halim, 2012; Hashim và cộng sự, 2014; Jayachandran và cộng sự, 2014). Cùng với nhau, tất cả các hợp chất chính này và một số chất chuyển hóa thơm dễ bay hơi có hàm lượng thấp tạo thành đặc tính có mùi thơm độc đáo của trầm hương.
Số lượng và loại thành phần chuyển hóa trầm hương của mỗi nghiên cứu được báo cáo khác nhau tùy thuộc vào nguồn trầm hương, phương pháp chiết xuất và phương pháp phân tích được sử dụng (Fazila và Halim, 2012; Jong và cộng sự, 2014; Pasaribu và cộng sự, 2015). Tuy nhiên, có hơn 150 hợp chất được xem xét bởi Naef (2011) đã được xác định cho đến nay trong trầm hương từ các nguồn khác nhau. Trong số các hợp chất này, có 70 sesquiterpen và khoảng 40 loại PEC đã được công nhận trong trầm hương và cấu trúc của chúng đã được làm sáng tỏ (Naef, 2011). Một số sesquiterpen đã được quan sát là có mặt thường xuyên hơn trong trầm hương từ các nghiên cứu khác nhau, bao gồm aromadendrene, agarospirol,-agarofuran, guaiol và (-)-aristolene (Fazila và Halim, 2012; Liu Y. và cộng sự, 2013; Jayachandran và cộng sự ., 2014; Jong và cộng sự, 2014; Hình 2). Một số sesquiterpen được báo cáo là đặc trưng cho loài, chẳng hạn như jinkoh-eremol và epi-γ-eudesmol chỉ có ở A. malaccensis, trong khi baimuxinal chỉ có ở A. crassna và A. sinensis (Naef, 2011; Liu Y. et cộng sự, 2013; Jong và cộng sự, 2014; Hashim và cộng sự, 2016). Điều đáng nói là trong nghiên cứu của Pasaribu et al. (2015), hàm lượng aromadendrene được tìm thấy nhiều hơn trong trầm hương cấp cao hơn và do đó nó được đề xuất như một chất đánh dấu hóa học hiệu quả để phân loại trầm hương. Bên cạnh aromadendrene, Jayachandran et al. (2014) sau đó đã đề xuất một hóa trị đánh dấu bổ sung có thể quan trọng trong việc phân loại dầu trầm hương.
Các chất dẫn xuất PEC, cũng như các thành phần hương thơm chính khác của trầm hương, là những thành phần quan trọng tạo nên hương thơm ngọt ngào, trái cây và lâu dài của trầm hương khi đốt. Các hợp chất này chỉ có thể được phát hiện bằng phương pháp chiết xuất bằng carbon dioxide và dung môi siêu tới hạn chứ không bao giờ có mặt trong dịch chiết của quá trình chưng cất thủy phân (Yoswathana, 2013; Jong et al., 2014). So với các thành phần sesquiterpene trong trầm hương, các loại PEC được xác định bởi GC-MS tương đối hạn chế. Các nghiên cứu về cấu trúc tiết lộ rằng tất cả các PEC được báo cáo trước đây trong trầm hương đều sở hữu cùng một khung cơ bản (trọng lượng phân tử: 250) và các nhóm thế tương tự, tức là các nhóm hydroxy hoặc methoxy (Mei và cộng sự, 2013). Tỷ lệ 2(2-phenylethyl) chromone và 2-(2-4-methoxy-phenylethyl) chromone trong trầm hương cao cấp như kanankoh có thể cao tới 66,47 %, cao hơn hẳn so với trầm hương chất lượng thấp hơn jinkoh mà chỉ có 1,5% (Ishihara et al., 1993). Hơn nữa, sự hiện diện của một số dẫn xuất PEC trong trầm hương được cho là hữu ích trong việc đánh giá phân loại các sản phẩm trầm hương (Shimada et al., 1982). Có 17 loại dẫn xuất chromone đặc trưng cho trầm hương và là chất đánh dấu tiềm năng cho mục đích xác thực (Naef, 2011). Các chromone được thay thế, chẳng hạn như agarotetrol và isoagarotetrol (Hình 3), đã được chứng minh là có mối tương quan tích cực với chất lượng của trầm hương thu được trên thị trường với một số ngoại lệ (Shimada et al., 1986).
Các loại và dẫn xuất của các hợp chất chính trong trầm hương rất rộng và đa dạng, cho thấy các đặc tính hương thơm khác nhau của trầm hương từ các loài và nguồn khu vực khác nhau. Cái nhìn sâu sắc hơn về các chất chuyển hóa của trầm hương chắc chắn sẽ tạo điều kiện thuận lợi cho việc xác định các dấu ấn sinh học được chấp nhận rộng rãi để phân loại trầm hương. Kể từ khi công bố đánh giá toàn diện của Naef (2011) về các thành phần chính của trầm hương, các hợp chất mới tiếp tục được phát hiện trong các nghiên cứu sau này (Wu và cộng sự, 2012a; Yang và cộng sự, 2014b; Wang và cộng sự, 2015 ). Số lượng các hợp chất được phát hiện trong trầm hương chắc chắn sẽ còn tăng lên trong tương lai.
Con Đường Sinh Tổng Hợp Các Thành Phần Trầm Hương
Sự hình thành trầm hương có thể liên quan đến cơ chế tự bảo vệ của cây Aquilaria trước các áp lực sinh học và phi sinh học (Gao et al., 2012b; Singh và Sharma, 2015). Căng thẳng kích hoạt các phản ứng phòng thủ của các loài Aquilaria, từ đó bắt đầu quá trình sinh tổng hợp chất chuyển hóa thứ cấp và tích tụ nhựa trầm hương. Trước đây, chúng tôi đã đề cập rằng các dẫn xuất sesquiterpenes và PEC là thành phần chính trong trầm hương. Do đó, điều cực kỳ quan trọng là phải hiểu con đường trao đổi chất để điều hòa và sinh tổng hợp sesquiterpene và các dẫn xuất chromone ở các loài Aquilaria để tạo ra sự hình thành trầm hương một cách hiệu quả.
Ở thực vật, các tiền chất isoprenoid cho quá trình sinh tổng hợp sesquiterpen, triterpen và sterol thường được cho là được cung cấp từ con đường axit mevalonic (MVA) trong tế bào chất. Trong plastid, con đường 1-deoxy-D-xylulose-5-phosphate (DXP) hay được gọi là methylerythritol phosphate (MEP) cung cấp tiền chất để sản xuất monoterpenes, diterpenes và carotenoids (Rohmer, 1999; Dong et al., 2015 ; Singh và Sharma, 2015). Hai con đường này sinh tổng hợp tiền chất isoprenoid đồng đẳng C5, đó là isopentenyl pyrophosphate (IPP) và đồng phân allylic điện di của nó dimethylallyl pyrophosphate (DMAPP). Một sự trao đổi IPP và DMAPP đã được quan sát xảy ra ở giữa plastid và cytosol ngay cả với sự phân chia không gian của hai con đường (Dong et al., 2015). Quá trình sản xuất tiền chất IPP và DMAPP từ pyruvate và acetyl-CoA liên quan đến một loạt enzyme theo con đường tương ứng (Hình 4). Các gen mã hóa cho các enzym này đã được xác định từ các loài Aquilaria thông qua phân tích trình tự phiên mã (Xu et al., 2013; Ye et al., 2016). Các đơn vị isoprene C5 này sau đó sẽ được đưa vào quá trình tạo ra C15 farnesyl pyrophosphate (FPP) bằng các phản ứng ngưng tụ tuần tự với sự có mặt của FPP synthase (FPS) (Rohmer, 1999; Yang và cộng sự, 2013; Ye và cộng sự, 2016) . FPS là một trong những enzyme giới hạn chính chịu trách nhiệm cho quá trình sinh tổng hợp sesquiterpene (Gaffe et al., 2000; Yang et al., 2013; Liu X. M. et al., 2017). Các gen mã hóa cho FPS đã được nhân bản từ Aquilaria microcarpa (Am-FaPS-1) (Kenmotsu và cộng sự, 2011) và Aquilaria sinensis (AsFPS1) (Yang và cộng sự, 2013). Mức độ phiên mã của AsFPS1 được báo cáo là cao hơn ở thân và rễ so với lá, cho thấy rằng quá trình tổng hợp sesquiterpene ở các loài Aquilaria có xu hướng đặc hiệu theo mô. Ngoài ra, biểu hiện của Am-FaPS-1 đã được chứng minh là được điều chỉnh tăng khi tiếp xúc với methyl jasmonate (MeJA), chiết xuất men và Ca2+-ionophore A23187, cho thấy rằng hai hóa chất trước đây có hiệu quả để bắt đầu quá trình sinh tổng hợp sesquiterpene trong khi Ca2+ có thể đóng vai trò là phân tử tín hiệu trong quá trình kích hoạt (Kenmotsu et al., 2011). Điều này cung cấp manh mối cho việc tạo ra trầm hương nhân tạo thông qua các phương pháp tiếp cận gây ra bởi hóa chất ngoại sinh bằng cách kích hoạt con đường sinh tổng hợp sesquiterpene trong cây Aquilaria.
Trong giai đoạn cuối cùng của quá trình sản xuất sesquiterpene, các enzyme chịu trách nhiệm cho sự đa dạng hóa sesquiterpene chủ yếu đến từ các lớp tổng hợp sesquiterpene (SesTPs) và mono-oxyase phụ thuộc cytochrom P450 (P450s). Các enzym SesTP chịu trách nhiệm xúc tác cho sự hình thành các phức hợp giàn giáo đa vòng từ FPP, sau đó là quá trình oxy hóa chức năng của các giàn giáo thu được bằng enzym cytochrom P450. Các nhóm hydroxyl được thêm vào bởi P450s có thể đóng vai trò xử lý phân tử để sửa đổi thêm, chẳng hạn như alkyl hóa, este hóa và bổ sung dư lượng đường (Pateraki et al., 2015). Ngoài ra, các enzyme P450 thực hiện quá trình hydroxyl hóa đặc hiệu lập thể trên các xương sống hydrocarbon, điều quan trọng đối với các chirality mới và các biến đổi tiếp theo của các phân tử sesquiterpene, cho đến nay chưa bao giờ được báo cáo từ các loài Aquilaria. Tương tự như vậy, các enzym oxyoreductase cytochrom P450 phụ thuộc NADPH (POR) trong Aquilaria đóng vai trò là đối tác oxy hóa khử của hoạt động xúc tác P450 về cơ bản chưa được khám phá. Một số nghiên cứu đã được báo cáo để phân lập các gen mã hóa cho SesTP từ Aquilaria, đây có thể được coi là những nỗ lực ban đầu để nghiên cứu SesTP liên quan đến sự hình thành trầm hương (Kumeta và Ito, 2010; Xu và cộng sự, 2013). Trong nghiên cứu của Kumeta và Ito (2010), năm gen mã hóa cho tổng hợp sesquiterpene chia sẻ trình tự axit amin rất giống nhau đã được phân lập từ Aquilaria crassna. Ba trong số các gen này đã được biểu hiện thành công ở Escherichia coli và FPP được chuyển đổi bằng enzym thành δ-guaiene làm sản phẩm chính của chúng. Ngoài ra, ba gen tổng hợp sesquiterpene khác (ASS1, ASS2 và ASS3) được xác định từ A. sinensis thông qua giải trình tự phiên mã đã được tiết lộ để mã hóa các enzyme cũng tạo ra δ-guaiene (Xu et al., 2013). Sự phân lập các gen SesTP cũng được mô tả ở A. malaccensis trong đó sự biểu hiện theo thời gian và không gian của hai SesTP được báo cáo trong nghiên cứu, tức là guaiene (AmGuaiS1) và sesquiterpene synthase (AmSesTPS1), đã được làm sáng tỏ (Azzarina và cộng sự, 2016) ). AmSesTPS1 được phát hiện có biểu hiện cao sau 6 giờ bị thương trong khi AmGuaiS1 được tạo ra sau 2 giờ bị thương ở cường độ lần lượt cao hơn 18 và 5,5 lần so với điều khiển không có vết thương. Gần đây, một gen tổng hợp sesquiterpene mới (As-sesTPS) đã được phân lập từ A. sinensis trong đó As-sesTPS tái tổ hợp đã xúc tác FPP thành nerolidol (Ye et al., 2018). Phân tích biểu hiện cho thấy mức độ phiên mã của As-sesTPS trong trầm hương cao hơn nhiều so với gỗ khỏe mạnh, ngụ ý rằng gen này có thể tham gia vào quá trình hình thành trầm hương. Mặc dù thực tế là nhiều hợp chất sesquiterpene đã được phát hiện từ trầm hương, nhưng các SesTP tương ứng chịu trách nhiệm sản xuất chúng vẫn chưa được báo cáo từ Aquilaria. Vì lý do đó, các con đường sinh tổng hợp sesquiterpene liên quan đến chức năng hóa terpene trong Aquilaria được khuyến khích để làm rõ thêm.
Mặt khác, chromone là một nhóm lớn các chất chuyển hóa thứ cấp với nhiều chỉ định điều trị tiềm năng đối với điều hòa miễn dịch, viêm, ung thư, tiểu đường, bệnh thần kinh, nhiễm trùng do vi khuẩn và virus (Khadem và Marles, 2011; Yang và cộng sự, 2012; Tawfik và cộng sự, 2014). Chromone có nguồn gốc từ một hợp chất hữu cơ đa vòng có tên là vòng benzopyran, với sự thay thế nhóm keto trên vòng oxime của nó. Người ta thường tin rằng các dẫn xuất của nhiễm sắc thể diễn ra do sự hội tụ của nhiều con đường sinh tổng hợp chất chuyển hóa thứ cấp liên quan đến con đường pentaketide, con đường axit shikimic và việc bổ sung phân tử nitơ từ axit amin hoặc các nguồn khác (Khadem và Marles, 2011). Do các đặc tính dược lý mở rộng liên quan đến cấu trúc vòng hai vòng của nó, các nhiễm sắc thể đã được sử dụng làm giàn giáo đặc quyền trong việc phát triển các loại thuốc mới (Reis et al., 2017). PEC là một loại nhiễm sắc thể nhỏ, chứa nhóm thế phenylethyl ở C2 của vòng benzopyran của nhiễm sắc thể có cấu trúc độc nhất trong họ (Ibrahim và Mohamed, 2015). Cho đến nay, PECs mới chỉ được phát hiện có mặt ở một số loài thực vật như Bothriochloa ischaemum (Wang et al., 2001), Imperata hình trụ (Liu X. et al., 2013), Cucumis melo L. (Ibrahim , 2014), Gyrinops salicifolia (Shao và cộng sự, 2016), và loài Aquilaria (Wu và cộng sự, 2012b; Yang và cộng sự, 2014a). Gần đây, một sơ đồ giả thuyết về con đường sinh tổng hợp của PEC đã được đề xuất bởi Liao et al. (2018) dựa trên phân tích chuyên sâu các thành phần hóa học của trầm hương bằng phương pháp GC-EL-MS và UPLC-ESI-MS/MS. Trong nghiên cứu của họ, PEC được phát hiện là thành phần chính của nhựa trầm hương, bao gồm chủ yếu là các nhiễm sắc thể 2-(2-phenylethyl) loại flindersia (FTPEC). Sự hình thành FTPEC được làm sáng tỏ thêm là có thể được xúc tác bởi polyketide synthase (PK) loại III thông qua quá trình ngưng tụ các chất tương tự dihydro-cinnamoyl-CoA và malonyl-CoA với 2-hydroxy-benzoyl-CoA để tạo ra giàn giáo PEC mà sau đó sẽ được xúc tác bởi hydroxylase hoặc O-methyltransferase (OMT) để tạo thành các FTPEC đa dạng về cấu trúc (Liao et al., 2018). Nghiên cứu gần đây cho thấy rằng áp lực về độ mặn có thể tạo ra quá trình sinh tổng hợp PEC ở A. sinensis calli (Wang và cộng sự, 2016). Phân tích phiên mã của các mô sẹo A. sinensis do muối này đã xác định được một số gen ứng cử viên được điều hòa tăng có khả năng tham gia vào quá trình sinh tổng hợp PEC, bao gồm ba gen mã hóa OMT (flavonol-OMT 1, flavonol-3-OMT và caffeoyl-CoA-OMT) và một gen tổng hợp polyketide loại III mã hóa cho chalcone synthase 1 (AsCHS1).
Bất chấp những tiến bộ gần đây đạt được dựa trên sự hiểu biết về quá trình sinh tổng hợp PEC, một nỗ lực to lớn là cần thiết để xác định bằng thực nghiệm các bước còn thiếu trong con đường sinh tổng hợp PEC phức tạp này. Ngay cả với kiến thức không đầy đủ về con đường sinh tổng hợp PEC chi tiết, việc tổng hợp nhân tạo các nhiễm sắc thể và một số dẫn xuất của nó vẫn khả thi do sự tiến bộ của các quá trình hóa học (Goel và Makrandi, 2006; Tawfik et al., 2014). Trầm hương là một nguồn dẫn xuất PEC phong phú, xứng đáng được nghiên cứu thêm để khám phá cấu trúc của các hợp chất chromone mới và cải thiện sự hiểu biết về cơ chế sinh tổng hợp của nó ở cấp độ phân tử.
Tín Hiệu Và Cơ Chế Điều Chỉnh Sự Hình Thành Trầm Hương
Trong tự nhiên, xác suất thu được cây Aquilaria chứa trầm hương là cực kỳ thấp (1–2%), chỉ có thể tìm thấy trên những cây bị nhiễm bệnh hoặc bị thương (Cui et al., 2013; Chhipa và Kaushik, 2017). Do đó, thật hợp lý khi cho rằng có một quá trình truyền tín hiệu cảm ứng vết thương gây ra sự biểu hiện của các chất tổng hợp sesquiterpene trước khi hình thành trầm hương. Để làm rõ mối quan hệ giữa truyền tín hiệu vết thương và điều hòa quá trình hình thành trầm hương, các nghiên cứu công suất cao về quá trình hình thành trầm hương gần đây đã được chú ý trong các nghiên cứu (Bảng 2). Sơ đồ cơ chế truyền tín hiệu được đề xuất của quá trình sinh tổng hợp và điều hòa sesquiterpene ở các loài Aquilaria được cung cấp trong Hình 4.
Con đường truyền tín hiệu protein kinase (MAPK) được kích hoạt bằng mitogen đã được đề xuất như là cơ chế truyền tín hiệu do vết thương gây ra cho sự hình thành trầm hương ở A. sinensis, mà phosphoryl hóa các yếu tố phiên mã xuôi dòng (TF) như MYB hoặc WRKY mà cuối cùng dẫn đến biểu hiện của sesquiterpene synthase gen (ASS) (Xu và cộng sự, 2013). Tầng tín hiệu MAPK bao gồm ba thành phần được kích hoạt tuần tự [MAPK kinase kinase (MAPKKKs), MAPK kinase (MAPKKs) và MAPKs], đây là cơ chế truyền tín hiệu được bảo tồn cao ở sinh vật nhân thực trong việc trung gian truyền tín hiệu ngoại bào đến các gen phản ứng xuôi dòng (Sinha et al., 2011; Xu C. và cộng sự, 2017). Xu et al. (2013) đã báo cáo rằng có tổng cộng 41 gen bản địa từ phân tích phiên mã của A. sinensis bị thương được chú thích là có liên quan đến con đường truyền tín hiệu MAPK và 25 con đường truyền tín hiệu canxi có thể đóng vai trò trong sự hình thành trầm hương do vết thương gây ra. Ở thực vật, các ion canxi (Ca2+) là các phân tử truyền tin thứ cấp nội bào quan trọng để điều chỉnh nhiều đường dẫn truyền tín hiệu phản ứng với các kích thích bên ngoài (Tuteja và Mahajan, 2007). Các nghiên cứu trước đây ở các loài thực vật khác đã chỉ ra rằng TF là bộ điều chỉnh quan trọng trong các đường truyền tín hiệu đáp ứng với căng thẳng để truyền tín hiệu đến các trung tâm tế bào khác nhau nhằm kích hoạt cơ chế thích ứng/bảo vệ thực vật chống lại môi trường bất lợi, bao gồm các TF như bZIP, ERF, EIN3, MYB, MYC và WRKY (Ambawat và cộng sự, 2013; Phukan và cộng sự, 2016; Schmiesing và cộng sự, 2016). Sự biểu hiện quá mức của AaWRKY1 trong Artemisia annua đã được phát hiện là đã điều chỉnh tích cực sự biểu hiện của gen tổng hợp amorpha-4,11-diene (ADS) và làm tăng đáng kể việc sản xuất artemisinin (Ma et al., 2009). Sự biểu hiện của bông TF GaWRKY cũng được chứng minh là điều chỉnh tăng các gen tổng hợp sesquiterpene cho quá trình sinh tổng hợp (+)-δ-cadinene và gossypol sesquiterpene (Xu et al., 2004).
Bên cạnh việc làm tổn thương cơ học, MeJA là một chất kích thích hiệu quả để tăng hàm lượng sesquiterpenes trong Aquilaria (Xu et al., 2013, 2016; Xu Y. H. et al., 2017). Các nghiên cứu trước đây đã chỉ ra rằng sốc nhiệt có thể làm tăng sự biểu hiện của các gen liên quan đến quá trình sinh tổng hợp axit Jasmonic (JA), bao gồm allene oxide cyclase (AOC), allene oxide synthase (AOS), lipoxygenase (LOX) và 12-oxophytodienoate reductase 3 (OPR3) ) gen, sau đó dẫn đến việc sản xuất JA và tích lũy sesquiterpene trong nuôi cấy tế bào huyền phù A. sinensis (Xu et al., 2016). Protein không nhạy cảm với coronatine A. sinensis 1 (AsCOI1), hoạt động như một thụ thể trong đường truyền tín hiệu MeJA, đã được nhân bản và đặc trưng (Liao et al., 2015). Nghiên cứu biểu hiện của AsCOI1 đã chứng minh rằng gen này được biểu hiện theo một kiểu mô cụ thể, cao nhất ở thân, tiếp theo là rễ và lá. Các phát hiện ngụ ý rằng việc sản xuất nhựa do các kích thích bên ngoài có thể phản ứng nhanh hơn trong thân cây Aquilaria. Gen AsCOI1 phản ứng nhanh với việc điều trị sớm MeJA, vết thương cơ học và stress nhiệt. Việc áp dụng MeJA ở A. sinensis đã làm tăng mức độ biểu hiện của 17 gen liên quan đến tín hiệu vết thương, bao gồm TF WRKY4 và MYB4, protein kinase CAPK, MAPK và MAPKK, NADPH oxidase noxB và một số bộ điều chỉnh liên quan đến phân tử tín hiệu MeJA, ethylene và hydro peroxide (Xu và cộng sự, 2013). Thật thú vị, hydro peroxide (H2O2) tạo ra NADPH oxidase noxB đã được phát hiện là được điều chỉnh tăng đáng kể bằng cách xử lý MeJA (Xu và cộng sự, 2013; Gong và cộng sự, 2017), phù hợp với những phát hiện rằng MeJA kích hoạt H2O2 sản xuất trong nhà máy (Orozco-Cardenas et al., 2001; Hung et al., 2006). Một nghiên cứu về nuôi cấy huyền phù A. sinensis cho thấy H2O2 có thể gây ra sự chết tế bào theo chương trình (PCD) và tổng hợp sesquiterpene bằng sự biểu hiện gia tăng của gen ASS do sự tích tụ nội sinh của axit salicylic (SA) (Liu và cộng sự, 2015) . Trong khi đó, sự biểu hiện của sesquiterpene synthase chính đáp ứng jasmonate ASS1 ở A. sinensis được mô tả là được điều chỉnh bởi TF AsMYC2 (Xu Y. H. et al., 2017). Là một gen đáp ứng ngay lập tức sớm đối với điều trị bằng MeJA, AsMYC2 liên kết với trình khởi động ASS1 có chứa mô-đun hộp G để bắt đầu biểu hiện ASS1. Tương tự như vậy, cấu trúc tương đồng của AsMYC2 trong Arabidopsis (MYC2) đã được chứng minh là phản ứng với MeJA và điều chỉnh tăng biểu hiện của hai chất tổng hợp sesquiterpene (TPS11 và TPS21) khi xử lý (Hong và cộng sự, 2012). Ngoài ra, xử lý bằng MeJA đã tạo ra thành công quá trình tổng hợp 3 sesquiterpen, đó là α-guaiene, α-humulene và δ-guaiene trong nuôi cấy tế bào A. crassna (Ito et al., 2005; Kumeta và Ito, 2010).
Trái ngược với con đường sinh tổng hợp sesquiterpene, quá trình sinh tổng hợp và điều hòa PEC hầu như vẫn chưa được biết đến. Trong nghiên cứu của Wang et al. (2016), 41 PEC đã được sản xuất trong A. sinensis calli bằng cách xử lý muối. Với sự trợ giúp của phân tích bản phiên mã thông lượng cao, đã thu được tổng cộng 18.069 bản phiên mã được thể hiện khác nhau giữa đối chứng và calli A. sinensis được xử lý bằng NaCl gây ra bởi áp lực độ mặn trong 24 hoặc 120 giờ. Nhiều gen biểu hiện khác nhau được báo cáo là có liên quan đến quá trình truyền tín hiệu hormone; bao gồm các gen mã hóa cho các tầng MAPK, kinase giống như thụ thể, các yếu tố phiên mã và truyền tín hiệu Ca2+ (Wang và cộng sự, 2016).
Tuy nhiên, tốc độ nghiên cứu không chỉ dừng lại ở việc tìm kiếm các chất cảm ứng hiệu quả và quá trình truyền tín hiệu cơ bản của chúng, mà còn mở rộng sang cơ chế điều hòa sau phiên mã của chúng. Bằng cách giải trình tự sâu các sRNA từ các mẫu A. sinensis kiểm soát khỏe mạnh và bị thương, Gao et al. (2012a) đã xác định được 10 miRNA phản ứng với áp lực từ 74 miRNA giả định được bảo tồn và các tiền chất hình thành kẹp tóc của chúng cũng đã được xác nhận. Mẫu biểu hiện tiết lộ rằng sáu trong số các miRNA phản ứng với căng thẳng này đã được điều chỉnh tăng, bao gồm miR159, miR168, miR171, miR396, miR397 và miR408, trong khi miR160 và miR398 được điều chỉnh giảm và tiếp tục ở mức giảm sau 2 ngày (Gao et al., 2012a). Các phản ứng khác nhau của miRNA đã phản ứng với việc điều trị và hiệu quả kéo dài trong các khoảng thời gian khác nhau phản ánh sự đa dạng về vị trí của chúng trong quá trình điều hòa sau phiên mã của phản ứng vết thương ở A. sinensis. Trong số các miRNA đã được xác định, miR398 được điều chỉnh giảm được quan tâm khi nó được chứng minh là điều chỉnh tiêu cực mô hình phân tử liên quan đến mầm bệnh (PAMP) do sự lắng đọng mô sẹo và khả năng miễn dịch bẩm sinh của thực vật chống lại vi khuẩn (Li et al., 2010). Phản ứng trái ngược của miR160 và miR398 ở A. sinensis gợi ý rằng chúng có thể là những chất điều chỉnh quan trọng và đóng một vai trò khác biệt hơn đối với sự hình thành trầm hương.
Nghiên cứu sâu hơn về hồ sơ miRNA của A. sinensis bị thương cho thấy rằng một số miRNA được bảo tồn tốt nhất như họ miR159 và miR396 đã tăng lên và giảm xuống nhanh chóng trong giai đoạn đầu điều trị cho thấy chức năng của chúng ở đầu nguồn của phản ứng vết thương (Gao Z. H. và cộng sự, 2014 ). MiR396b2 được điều chỉnh giảm trong các mô bị thương của A. sinensis được cho là có liên quan đến quá trình sinh tổng hợp và tích lũy các thành phần của trầm hương. Đích của miR396b2, có hoạt tính glutamyl-tRNA reductase, được cho là sẽ liên kết NADP và tạo ra NADPH. NADPH là đồng yếu tố cho hai enzyme chủ chốt trong quá trình sinh tổng hợp terpene, đó là 1-deoxy-D-xylulose 5 phosphate (DXR) và 3-hydroxy-3-methylglutaryl-CoA (HMGR) (Nagegowda, 2010). Hơn nữa, các P450 thực vật có chức năng oxy hóa các giàn giáo terpene cũng yêu cầu các chất khử cho hoạt động xúc tác của nó, thường được cung cấp bởi NADH hoặc NADPH. Mặc dù trong tình trạng thiếu bộ gen được giải trình tự, phân tích phiên mã thông lượng cao cung cấp một phương pháp khả thi để kiểm tra những thay đổi tổng thể của biểu hiện gen trên các loài Aquilaria phản ứng với nhiều loại căng thẳng. Nghiên cứu sâu hơn về chức năng của các trình tự điều chỉnh đã xác định sẽ giúp tiết lộ cơ chế điều hòa của sự hình thành trầm hương.
Triển vọng tương lai của công nghệ cảm ứng trầm hương
Các nghiên cứu trước đây đã chỉ ra rằng sự hình thành trầm hương có thể bị ảnh hưởng bởi nhiều yếu tố. Cùng với thành phần nhựa trầm hương cực kỳ phức tạp, người ta tin rằng sự hình thành trầm hương là một quá trình phức tạp liên quan đến nhiều thay đổi sinh lý xảy ra trên cây Aquilaria để đối phó với các kích thích bên ngoài dưới dạng sinh học hoặc phi sinh học. Toàn bộ quá trình hình thành trầm hương này trong mọi trường hợp không thể tách rời khỏi phản ứng biểu hiện gen của cây đối với các yếu tố kích hoạt. Do đó, việc cải thiện công nghệ cảm ứng trầm hương trong tương lai cần nhấn mạnh vào hai khía cạnh là nâng cao hơn nữa hiệu quả cảm ứng và sàng lọc các dòng Aquilaria đáp ứng tốt hơn cho sản xuất nhựa theo chương trình nhân giống.
Để nâng cao hiệu quả cảm ứng, kỹ thuật cảm ứng đóng vai trò quyết định. Khái niệm về các phương pháp cảm ứng hiện có có thể được tóm tắt là cung cấp các kích thích bên ngoài để kích hoạt quá trình sản xuất các phân tử tín hiệu thực vật mà cuối cùng dẫn đến quá trình sinh tổng hợp nhựa hoặc bỏ qua các kích thích bên ngoài thông qua việc đưa trực tiếp các phân tử tín hiệu vào thực vật. Trong mọi trường hợp, mối quan tâm chung là tăng sản lượng và chất lượng trầm hương cũng như giảm sự can thiệp của con người (ví dụ: quy trình đục lỗ) trong quá trình cảm ứng. Do phương pháp gây tổn thương vật lý và cảm ứng sinh học đã nói ở trên có những nhược điểm không thể tránh khỏi của chúng là chất lượng trầm hương không ổn định và đòi hỏi lực lượng lao động chuyên sâu, nên phương pháp cảm ứng hóa học có thể được coi là một phương pháp đầy hứa hẹn để tối ưu hóa hơn nữa (Bảng 1). Sự hiểu biết toàn diện về sự hình thành trầm hương ở cấp độ phân tử thông qua công suất cao sử dụng phương pháp tiếp cận omics như trancriptomic và metabolomic dường như có lợi cho việc cải thiện công thức cảm ứng có mục tiêu và định hướng hơn là các thử nghiệm dựa trên thử nghiệm và sai sót.
Sự tích hợp của các phương pháp sinh học hệ thống và omics, bao gồm bộ gen, phiên mã, proteomics, chuyển hóa và phân tích chức năng; cung cấp một giải pháp tiềm năng để hiểu được bản chất đa gen của quá trình sinh tổng hợp nhựa ở Aquilaria. Trên cơ sở một số lượng lớn các thí nghiệm cảm ứng trầm hương đã được tiến hành trước đó, các nghiên cứu về phiên mã và các chất chuyển hóa (Naef, 2011; Gao X. và cộng sự, 2014; Ye và cộng sự, 2016; Wu và cộng sự, 2017), phân tích omics tích hợp có thể thực sự đóng vai trò là nền tảng để xây dựng một bức tranh toàn diện hơn về cơ chế sinh tổng hợp nhựa trầm hương liên quan đến các lớp omics khác nhau. Sự phát triển của phép đo khối phổ thông lượng cao (MS), microarray và công nghệ giải trình tự (DNA và RNA) đã cho phép tích hợp dữ liệu đó vào khung sinh học hệ thống thông qua omic tích hợp giúp dự đoán tương tác gen-gen, xác định trình điều khiển gen và các dấu hiệu phân tử của sự hình thành trầm hương (Hình 5). Tương tự như vậy, các phân tử tín hiệu tiềm năng cho sự hình thành trầm hương có thể được dự đoán sau khi dữ liệu sẵn có tăng lên đáng kể này. Các phân tử tín hiệu hiệu quả đã được xác thực bằng thực nghiệm có thể được thêm vào công thức hiện có của chất cảm ứng hóa học để nâng cao hơn nữa hiệu suất cảm ứng. Hơn nữa, việc xác định trước thành phần nhựa hoặc chất lượng trầm hương có thể hình dung được với sự hiểu biết sâu sắc về các chỉ số chất lượng chính và các quy trình cụ thể liên quan đến sản xuất trầm hương bằng cách làm sáng tỏ tích hợp các cấu hình chuyển hóa và phiên mã khác nhau theo các phương pháp cảm ứng khác nhau. Việc cải thiện hiệu quả cảm ứng trầm hương trong tương lai nên đi kèm với việc phát triển hệ thống giám sát để phát hiện sớm những cây không đáp ứng để có thể tránh chặt hạ những cây Aquilaria không được kích thích thành công. Điều này có thể đạt được bằng cách theo dõi sự biểu hiện của một bộ gen liên quan đến quá trình sinh tổng hợp nhựa trầm hương.
Bên cạnh chất cảm ứng hiệu quả, khả năng đáp ứng của cây Aquilaria đối với kích thích là một yếu tố quyết định khác để sản xuất trầm hương. Khi biết rằng mức độ phản ứng của thực vật đối với các tác nhân kích thích chủ yếu phụ thuộc vào cấu tạo di truyền của chúng, việc sử dụng dòng Aquilaria có tính phản ứng cao làm mục tiêu cảm ứng dự kiến sẽ làm tăng thêm sản lượng trầm hương thay vì chỉ tối ưu hóa công thức cảm ứng. Thông thường, nhân giống chọn lọc dựa trên chọn lọc kiểu hình đã được áp dụng để phát triển các dòng cây trồng mới với các tính trạng mong muốn. Công nghệ hiện tại liên kết những đặc điểm có lợi này của thực vật với các dấu hiệu di truyền (biến thể DNA/RNA) hoặc sinh hóa (chất chuyển hóa đặc trưng) để cho phép lựa chọn có sự hỗ trợ của dấu hiệu (MAS). Cách tiếp cận của MAS mang lại nhiều hứa hẹn cho việc lựa chọn các dòng Aquilaria ưu tú vì những dấu ấn sinh học này có thể được áp dụng để dự đoán các đặc điểm kiểu hình trước khi các đặc điểm này phát triển thành đáng chú ý hơn. Các dấu ấn sinh học như vậy cũng có thể được sử dụng để phát triển các xét nghiệm chẩn đoán nhanh và có mục tiêu sẽ hỗ trợ chương trình lựa chọn. Là một giải pháp thay thế để thu được dòng năng suất cao, phương pháp tiếp cận kết hợp kỹ thuật di truyền (ví dụ: công nghệ chỉnh sửa bộ gen CRISPR-Cas9) với nuôi cấy mô có thể tạo ra khả năng điều khiển các gen điều hòa chính của Aquilaria tham gia vào quá trình sản xuất trầm hương. để tinh chỉnh hoặc chuyển hướng dòng trao đổi chất theo con đường trao đổi chất mong muốn. Nhìn chung, một chiến lược tích hợp và thông lượng cao sẽ cung cấp đầy đủ thông tin để liên tục cải thiện các phương pháp cảm ứng trầm hương, vượt trội so với cách thiết lập phương pháp cảm ứng truyền thống dựa trên quan sát trực quan và kinh nghiệm cá nhân. Việc hiểu sâu hơn về các hợp chất thiết yếu và cơ chế sinh tổng hợp của nhựa trầm hương sẽ giúp dễ dàng kiểm soát sự ổn định về sản lượng, chất lượng và giá cả của trầm hương trong tương lai.
Từ vựng ngành trầm:
1. resinous part: 树脂部分 / phần nhựa
2. endangered: 濒危的 (Bīnwēi de) / bị đe dọa
3. Aquilaria tree: 沉香树 / cây dó bầu
4. physiologically triggered Aquilaria tree: 生理触发沉香树 / cây dó bầu kích thích sinh học
5. aromatic dark resinous heartwood: 芳香深色树脂心材 /tâm gỗ nhựa sẫm thơm
6. The formation of agarwood: 沉香的形成 / Sự hình thành trầm hương
7. associated with the wounding and fungal infection of the Aquilaria trees: 与沉香树的受伤和真菌(junf)感染有关 / liên quan đến vết thương và nhiễm nấm của cây Aquilaria
8. The resin is secreted by the trees as defense reaction and deposited around the wounds over the years following the injury, where the accumulation of the volatile compounds eventually forms agarwood: 树脂作为防御(fangsyuf)反应由树木分泌(mif),并在受伤后的数年内沉积在伤口周围,挥发性化合物的积累最终形成沉香 / Nhựa do cây tiết ra như một phản ứng tự vệ và lắng đọng xung quanh vết thương trong nhiều năm sau vết thương, nơi mà sự tích tụ của các hợp chất dễ bay hơi cuối cùng tạo thành trầm hương.
9. therapeutic perfumes: 治疗香水 / nước hoa trị liệu
10. aromatic food ingredient: 芳香食品成分 / thành phần thực phẩm taoj thơm
11. sedative: 镇静剂 / thuốc an thần
12. pain reliever: 去痛药 / thuốc giảm đau
13. digestive aid: 助消化 / hỗ trợ tiêu hóa
14. carminative: 驱风的 (qufeng) / tống hơi
15. agarwood essential oil: 沉香精油 / tinh dầu trầm hương
16. Thymelaeaceae / 瑞香科 (ruif) / chi dó trầm
17. angiosperms / 被子植物 /thực vật hạt kín
18. The destructive exploitation of agarwood: 沉香的破坏性开发 / Khai thác trầm hương một cách tận diệt
19. Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES): Công ước về buôn bán quốc tế các loài động, thực vật hoang dã nguy cấp (CITES) / 濒(bin危野生动植物种国际贸易公约(CITES)
20. drastic declination: 急剧偏(pian)角 / suy giảm nghiêm trọng
21. depletion: 消耗 / cạn kiệt
22. mass cultivation and large plantation of Aquilaria trees: 沉香树的大规模种植和大面积种植 / trồng ồ ạt và trồng đại trà cây dó bầu
23. scarcity: 缺乏 (que fas) sự khan hiếm
24. artificially induce the agarwood formation: 人工诱(youf)导沉香形成 / nhân tạo tạo ra trầm hương
25. resin deposition: 树脂沉积 / lắng đọng nhựa
26. internal tissues: 内部组织 / các mô bên trong
27. mechanical wounding approach: 机械致伤法 (ji xief shif) / phương pháp vết thương cơ học
28. pathogen-inducing techniques: 病原体诱(youf)导技术 / kỹ thuật gây bệnh
29. indiscriminate harvesting: 乱采 / thu hoạch bừa bãi
30. regeneration: 再生 / sự tái tạo
31. disease infestations: 疾病侵扰 / (jisbingf qin raor) dịch bệnh
32. pathogenic microbes: 病原微生物 / vi khuẩn gây bệnh
33. diverse artificial induction methods: 多种人工诱导方式 / phương pháp cảm ứng nhân tạo đa dạng
34. bark removal: 去除树皮 / loại bỏ vỏ cây
35. inferior quality: 劣质 (lief zhif) / dưới mức chuẩn
36.